
Le cerveau des requins et des chimères
Le cerveau est le principal organe du système nerveux des animaux. Chez les vertébrés (Figure 1), il est situé dans la tête et protégé par une boite crânienne cartilagineuse ou osseuse. Il constitue le centre de régulation des autres organes et le siège des fonctions cognitives.
Le cerveau des vertébrés à mâchoires (gnathostomes) est constitué, de l’avant vers l’arrière, de six régions principales : les bulbes olfactifs, le télencéphale, le diencéphale, le mésencéphale, le cervelet et la moelle allongée. Les premiers vertébrés à mâchoires, apparus il y a plus de 500 millions d’années, se différencient des vertébrés sans mâchoires par, entre autre, la présence de mâchoires et d’un cervelet. Ce dernier a un rôle central dans le contrôle moteur, permettant une diversité nouvelle de prises alimentaires et de prédations, et des modes de locomotion plus actifs. Le cervelet est donc une innovation évolutive majeure des gnathostomes.
Au cours de l’évolution des gnathostomes, le cerveau va se diversifier, notamment en modifiant la proportion relative de ses six régions, en fonction des relations de parenté (phylogénie), des différents environnements et des différents modes de vie de ces animaux.
C’est ce qui est présenté ici à travers l’exemple des chondrichthyens (« poissons cartilagineux ») tels que requins et chimères.
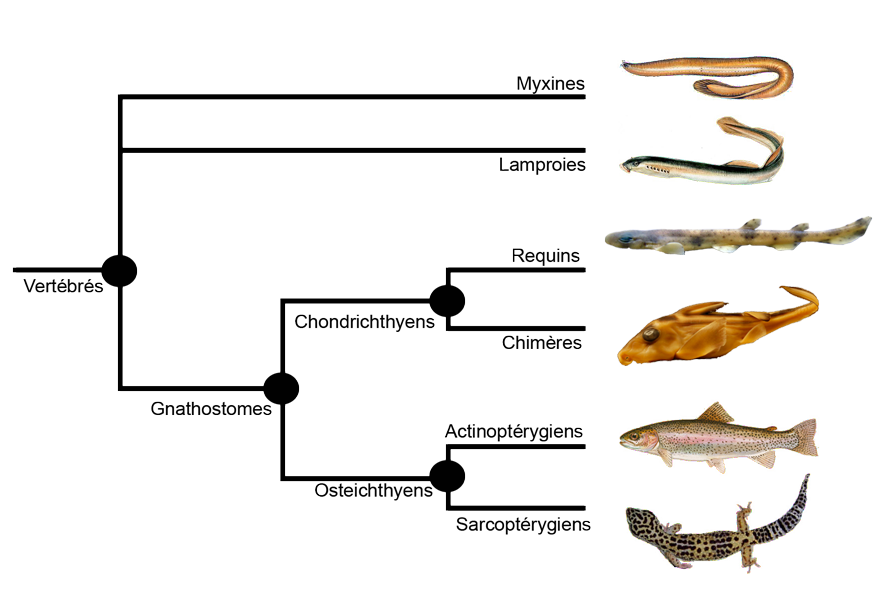
Figure 1. Arbre phylogénétique des vertébrés actuels
Les chondrichthyens sont apparus il y a plus de 400 millions d’années. Leur squelette n’est pas constitué de vrai os, mais de cartilage comme chez les poissons sans mâchoires actuels (lamproies, myxines). C’est en partie pour cette raison que les chondrichthyens ont été perçus comme des « reliques du passé », avec une morphologie primitive pour les gnathostomes, et n’ayant pas « évolués » depuis des centaines de millions d’années. Cette vision s’appliquait également à l’évolution du cerveau des chondrichthyens et les anciens auteurs considéraient que les requins devaient posséder un cerveau relativement petit et archaïque par rapport aux autres vertébrés à mâchoires.
L’essor récent des techniques de visualisation 3D a permis d’avoir une meilleure appréciation de l’anatomie et de l’évolution du système nerveux central car elles facilitent la visualisation de ces structures internes in situ, en position naturelle, et sans artéfact de préparation manuelle.
Ci-dessous sont reconstitués en trois dimensions les cerveaux de deux chondrichthyens actuels de plus en plus utilisés comme modèle biologique dans les sciences de l’évolution : la roussette Scyliorhinus canicula (Figure 2A), un petit requin commun de nos côtes, et la chimère Callorhinchus milii (Figure 2B), un holocéphale vivant au large de la Nouvelle Zélande. Les deux spécimens présentés ici sont des embryons au dernier stade de développement, c’est-à-dire juste avant leur éclosion.
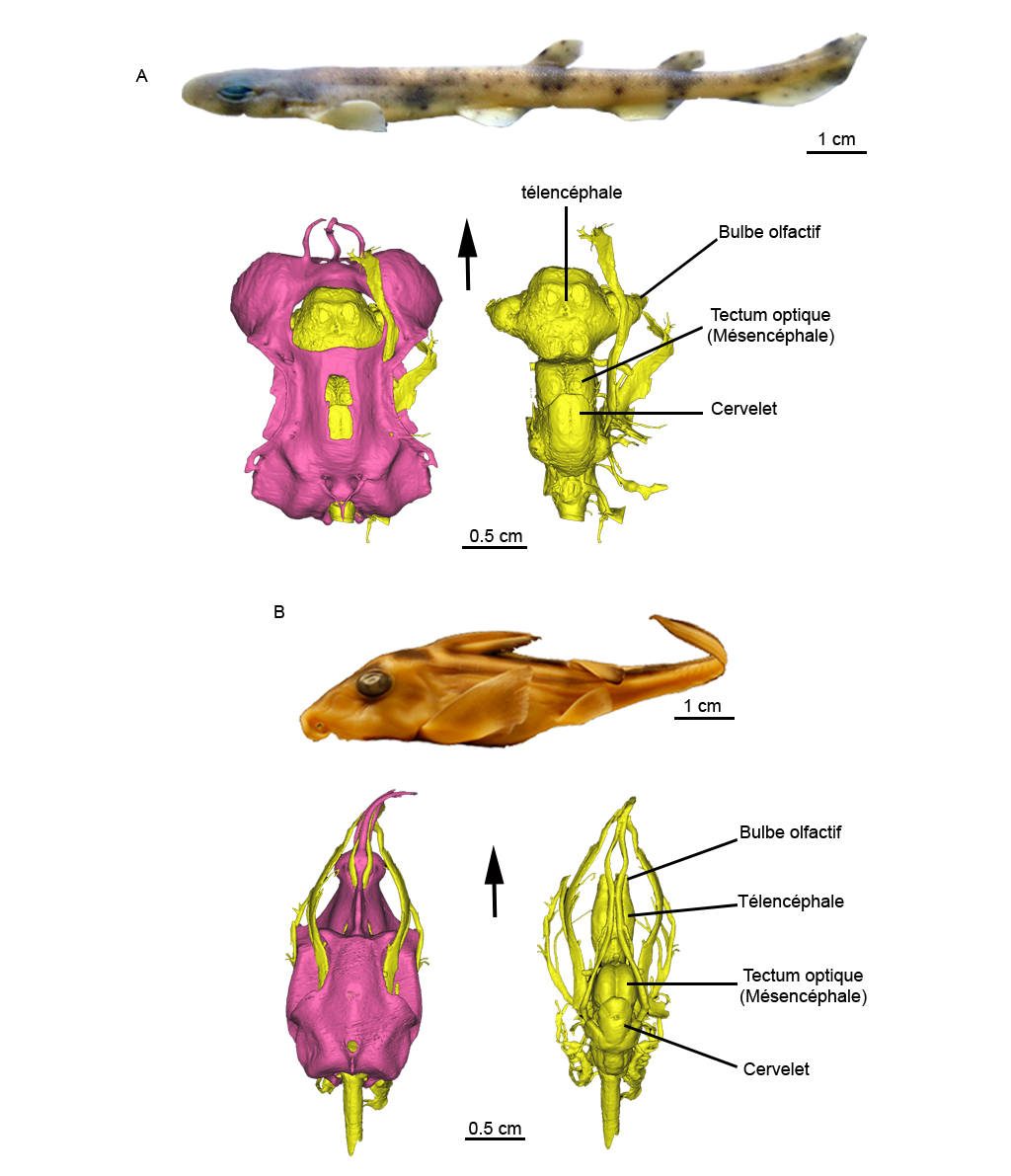
Figure 2. Reconstitution virtuelle à partir de données tomographiques par synchrotron (ESRF, Grenoble) de la petite roussette Scyliorhinus canicula (A) (en attente d'entrée dans les collections MNHN) et de la chimère Callorhinchus milii (B) (Collection Academy of Natural Sciences in Philadelphia, Philadelphia, Pennsylvannie, USA). Vue latérale gauche pour les photos et vue dorsale pour les reconstitutions 3D, les flèches indiquent l’avant ; rose : crâne ; jaune : cerveau.
L’idée ancienne qu’il existe une complexification croissante du cerveau des poissons jusqu’à l’homme ne supporte pas la réalité de l’observation : la diversité morphologique du cerveau des poissons est bien plus importante que celle des autres vertébrés. Ceci peut s’expliquer par un nombre important de stimuli sensoriels dans les environnements aquatiques. Ainsi, en plus de la vision, de l’odorat, du toucher, du goût et de l’ouïe, les poissons possèdent des “sens aquatiques” supplémentaires, comme la ligne latérale qui perçoit les vibrations de l’eau et du son, ou encore des systèmes détectant les champs électromagnétiques comme les ampoules de Lorenzini des chondrichthyens.
De plus, les chondrichthyens possèdent des stratégies reproductives, des comportements sociaux et des niches écologiques complexes et diversifiés qui façonnent la morphologie du cerveau. Ainsi, il existe dans ce groupe une diversité neuro-morphologique très importante.
Contrairement à ce que l’on pensait, le cerveau des chondrichthyens est aussi grand (en proportion de la taille corporelle totale) que celui des oiseaux et des mammifères. Chez les chondrichthyens, la taille relative du cerveau a principalement un signal écologique et l’augmentation de la taille se fait indépendamment dans chaque grande lignée. Les espèces récifales possèdent un cerveau relativement plus grand que les requins du grand large, avec un télencéphale et un tectum optique développés mais des bulbes olfactifs réduits. Ces caractéristiques reflètent des capacités cognitives plus importantes liées à la vie dans des organisations spatiales complexes que sont les récifs coralliens et des interactions sociales importantes.
Les espèces pélagiques et prédatrices actives ont un système visuel performant : le tectum optique est large, le cervelet imposant et complexe avec circonvolutions, et les bulbes olfactifs réduits. Dans le même ordre d’idée, les espèces à prédation passive ont généralement un cerveau plus petit, même si de notables exceptions existent, comme les grandes raies manta, planctonivores, qui possèdent un des cerveaux les plus gros des chondrichthyens.
Il est également intéressant de noter que, malgré une lointaine relation de parenté, il existe une même tendance de la croissance relative de quelques régions du cerveau chez les requins et chez les mammifères. Par exemple, le télencéphale et le cervelet, les régions responsables de la complexité comportementale et motrice, ont une taille qui a proportionnellement fortement augmenté au cours de l’évolution, chez les requins comme chez les mammifères.
Le cerveau des chondrichthyens peut également avoir un signal phylogénétique (c’est-à-dire qui permet de retracer les relations de parentés des organismes).
Par exemple, le cerveau des chimères a un long et étroit télencéphale chez l’adulte, alors qu’il est très court chez les requins. Pour certains auteurs cette structure rapprochait les chimères des ostéichthyens (poissons osseux et vertébrés terrestres). Le modèle 3D de l’embryon de la chimère montre qu’en réalité le nerf olfactif est court comme chez le requin. Chez l’adulte, le télencéphale entier est déplacé vers l’avant et un long pédoncule le sépare du reste du cerveau. Il s’agit ici d’une condition particulière du télencéphale bien différente de celle des osteichthyens.
Pour retrouver la forme ancestrale du cerveau des chondrichthyens et les modes de vie des requins du passé, il faudrait pouvoir avoir accès à des cerveaux fossilisés. Seulement cet organe, composé de tissus mous, ne se fossilise que très rarement. C’est pourquoi les études de paléoneurologie se sont basées sur des moulages de la cavité crânienne pour obtenir une approximation de la morphologie du cerveau qu’elle abritait. Si cela fonctionne assez bien pour les mammifères, chez qui le cerveau rempli cette cavité, ce n’est pas le cas pour l’immense majorité des poissons, chez qui le cerveau est considérablement plus petit que la cavité qui l’abrite. Chez certains requins adultes le cerveau n’occupe que 6% de la cavité !
Avec les nouvelles techniques 3D, et en particulier la tomographie à rayons X par lumière synchrotron, de plus en plus de tissus mous fossilisés sont découverts chez des fossiles. Ainsi, pour la première fois, un cerveau fossilisé a été mis au jour chez une chimère fossile vieille de 300 millions d’années (Figure 3). Ce crâne de chimère fossile, nommée Iniopera, a été découvert dans un nodule de pierre de quelques centimètres, dans des terrains carbonifères des Etats-Unis. Ce cerveau apparaît petit par rapport à la cavité crânienne dont la morphologie ne reflète pas exactement celle du cerveau. Le tectum optique est important et le cervelet relativement petit : cette chimère devait habiter un milieu peu profond, peut-être récifal, et devait être un prédateur relativement lent, comme les chimères actuelles.
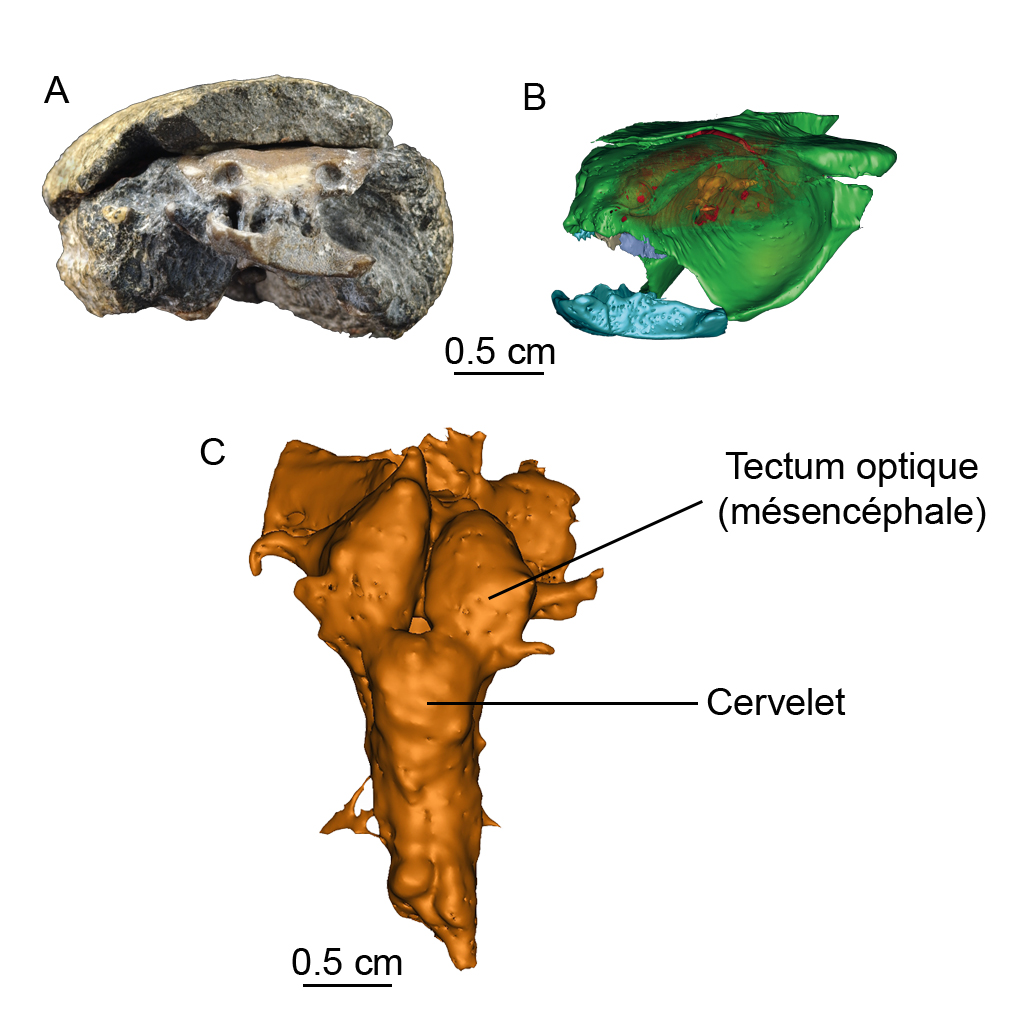
Figure 3. La chimère fossile Iniopera (collection Kansas University Natural History Museum, Lawrence, Kansas, USA, n°KUNHM 22060). A : nodule avec crâne fossile en vue frontale ; B : reconstitution virtuelle à partir de données tomographiques par synchrotron (ESRF, Grenoble) du crâne (vert), de la cavité crânienne (rouge) et du cerveau (orange) en vue oblique ; C : cerveau en vue dorsale. Ci-dessous animation du modèle 3D de ce spécimen fossile.
Pour conclure, l’étude du cerveau des chondrichthyens actuels et fossiles, facilitée par les nouvelles techniques de visualisation 3D, contredit la vision ancienne qui décrivait les chondrichthyens comme des organismes primitifs à morphologie simple et archaïque.
Alan Pradel - UMR 7207 - MNHN
Références